Каково строение и функции рибосом. Рибосомы — строение и функции. Расположение в клетке
Рибосомы (ribosome) является немембранные органелл клетки, состоящий из рРНК и рибосомных белков (протеинов). Рибосомы осуществляет биосинтез белков транслируя с мРНК полипептидную цепь. Таким образом, рибосому можно считать фабрикой, производящей белки, основываясь на имеющейся генетической информации. В клетке созревшие рибосомы находятся преимущественно в компартментах, для активного белкового синтеза. Они могут свободно плавать в цитоплазме или быть прикрепленными к цитоплазматического стороны мембран эндоплазматического ретикулума или ядра. Активные (те что есть в процессе трансляции) рибосомы находятся преимущественно в виде полисом. Существует ряд свидетельств, указывающих на то, что рибосома является рибозимов.
Исторический обзор
Рибосомы было обнаружено в начале 1950-х годов. Первое глубокое исследование и описание рибосом, как клеточных органелл, было совершено Джорджем Паладе (George E. Palade). По имени исследователя, рибосомы были названы «частицами Паладе», но впоследствии, в 1958 году, их было переименовано в «рибосомы», учитывая высокое содержание РНК. Роль рибосом в биосинтезе белков было установлено более десятилетием позже.
Синтетическая рибосома
После отсоединения от мРНК и началом нового раунда трансляции рибосомальные малая и большая субъединицы отделяются друг от друга. Поэтому, создание синтетической рибосомы было технически сложным, поскольку синтетические и имеющиеся в клетке субъединицы смешивались от раунда к раунду трансляции.
Начиная с конца 90-х годов 20 века удалось создать несколько видов мутантных малых субъединиц рибосомы, которые имели специфическую последовательность в 16S рРНК и соединялись с мРНК, в которой последовательность Шайна-Дальгарно была специфически синтезирована для взаимодействия с модифицированной 16S рРНК. Это позволило выполнять отбор мутировавших малых субъединиц РНК от нативных и интрудукуваты несколько мутаций для изучения свойств синтеза белка.
Однако большая рибосомальная субъединица представила проблемы, поскольку при создании синтетического варианта не бело возможности заставить его отделяться от мРНК или от малой субъединицы после завершения одного раунда трансляции. Большая субъединица содержит важные для изучения структуры, такие как канал для выхода синтезируемого белка и сайт PTC (англ. Peptidyl transferase centre), в котором происходит соединение аминокислоты, присоединена к тРНК, которая находятся на А-сайте рибосомы, к пептидильного цепи, который соединен к молекуле тРНК, которая находится на P-сайи рибосомы
В июле 2015 года удалось синтезировать первую полностью синтетическую рибосому. Для того, чтобы большая и малая субъединицы НЕ отсоединялись, их было связано в одну молекулу путем синтеза 16S-23S конструкта (Ribo-T). Такая синтетическая рибосома успешно выполняла синтез белка не только in vitro, но и поддерживала рост E.coli при вынужденной отсутствия нативных рибосом.
Строение рибосомы
Общее строение
Рибосомы прокариот и эукариот очень похожи по строению и функции, но отличаются размером. Они состоят из двух субъединиц: одной большой и одной малой. Для процесса трансляции необходимо слаженное взаимодействие обеих субъединиц, вместе составляют комплекс с молекулярной массой несколько миллионов Дальтон (Da). Субъединицы рибосом обычно обозначаются единицами Сведберга (S), является мерой скорости седиментации при центрифугирования и зависят от массы, размера и формы частицы. Обозначены в этих единицах, большая субъединица является 50S или 60S (прокариотические или эукариотические, соответственно), имела является 30S или 40S, и целая рибосома (комплекс малой вместе с большой) 70S или 80S.
Молекулярный состав
Молекулярный состав рибосом является достаточно сложным. Например, рибосома дрожжей "Saccharomyces cerevisiae" состоит из 79 рибосомных белков и 4 различных молекул рРНК. Биогенез рибосом также чрезвычайно сложным и многоступенчатым процессом, происходящим в ядре и ядрышке эукариотической клетки.
Атомная структура большой субъединицы (50S) организма Haloarcula marismortui была опубликована N. Ban et al. В журнале Science 11 августа 2000. Вскоре после этого, 21шого сентября 2000 года, BT Wimberly, et al., Опубликовали в журнале Nature структуру 30S субъединицы организма Thermus thermophilus. Используя эти координаты, MM Yusupov, et al. Сумели реконструировать целую 70S частичку Thermus thermophilus и опубликовать ее в журнале Science, в Мае 2001 В 2009 году профессор Джордж Чьорч (George Church) и коллеги из Гарварда создали полностью функциональную искусственную рибосому в обычных условиях, которые присутствуют в клеточном среде. Как конструкционные элементы использовались молекулы с расщепленной с помощью энзимов кишечной палочки. Созданная рибосома успешно синтезирует белок, отвечающий за биолюминесценцию.
Центры связывания РНК
Рибосомы содержит четыре сайты связывания для молекул РНК: один для мРНК и три для тРНК. Первый сайт связывания тРНК называется сайтом "аминоацил-тРНК ", или" А-сайтом ". В этом сайте содержится молекула тРНК "заряженная" "следующей" аминокислотой. Другой сайт, "пептидил-тРНК "связывающий, или" P-сайт ", содержит молекулу тРНК, связывает растущий конец полипептидной цепи. Третий сайт, это "сайт выхода", или "E-сайт". В этот сайт попадает пустая тРНК которая избавилась растущего конца полипептида, после его взаимодействия с последующей "заряженной" аминокислотой в пептидильному сайте. Сайт связывания мРНК находится в малой субъединицы. Он удерживает рибосому "нанизанной" на мРНК которую рибосома транслирует.
Функция
Рибосомы являются органелл, на которой происходит трансляция генетической информации, закодированной в мРНК. Эта информация воплощается в синтезированный тут же полипептидную цепь. Рибосомы несет двоякую функцию: является структурной платформой для процесса декодирования генетической информации с РНК, и владеет каталитическим центром ответственным за формирование пептидной связи, так называемым "пептидил-трансферазним центром". Считается пептидил-трансферазна активность ассоциируется с рРНК, и поэтому рибосома является рибозимов.
Локализация рибосом
Рибосомы классифицируются как свободные (находятся в гиалоплазме) и несвободные или прикрепленные (связанные с мембранами эндоплазматической сети).
Свободные и прикреплены рибосомы отличаются только локализацией, но они структурно идентичны. Рибосому называют свободной или прикрепленной в зависимости от того белок синтезируемый имеет ЭР-нацеленную сигнальную последовательность, поэтому индивидуальная рибосома может быть прикрепленной создавая один белок, но свободной в цитозоле когда создает другой белок.
Рибосомы иногда называют органеллами, но использование термина органеллы ограничивается субклеточном компонентами которых фосфолипидную мембрану, а рибосома (being entirely particulate) таковой не является. Поэтому рибосомы иногда описывают как "немембранные органеллы".
Общая информация
В эукариотических организмах рибосомы можно найти не только в цитоплазме, но и внутри в некоторых крупных мембранных органеллах, в частности в митохондриях и хлоропластах. Строение и молекулярный состав этих рибосом отличается от состава обще-клеточных рибосом, и является близким в состав рибосом прокариот. Такие рибосомы синтезируют органелл-специфические белки, транслируя органелл-специфическую мРНК.
В эукариотических клетках долгое время считалось, что рибосомы, прикрепленные к эндоплазматического ретикулума выполняют синтез белков, которые будут секретируемого наружу или трансмембранных или других сигнальных белков, присоединенных к плазмалеммы. Рибонуклеопротеин SRP (англ. Signal recognition particle) выполняет распознавание тех белков в процессе синтеза, которые должны быть трансмембранными и присоединяет рибосому к эндоплазматического ретикулума. Однако в последнее время исследования указывают, что 50-75% рибосом могут быть прикреплены к ЭПР за не до конца выяснены механизмы и большинство белков в клетке проходит синтез в рибосомах, прилегающих к ЭПР. Так, в клеточной линии HEK-293 75% мРНК видповидяе цитозольным белкам, однако до 50% рибосом связанные с ЭПР.
Болезни
Считается, что генетические дефекты рибосомных белков и факторов биогенеза рибосом является летальными на ранних эмбриональных стадиях развития высших организмов. Экспериментальный мутагенез рибосомных белков в Drosophila melanogaster (мутации minute) вызывает общий фенотип: заниженная скорость митоза, уменьшен размер тела, заниженная фертильность, короткие реснички. Существует ряд свидетельств связывающих раковую трансформацию клеток млекопитающих с расстройствами трансляционной системы в целом и расстройствами системы биогенеза рибосом в частности.
На заре развития жизни на Земле все клеточные формы были представлены бактериями. Они всасывали органические вещества, растворённые в первичном океане, через поверхность тела.
Со временем некоторые бактерии приспособились производить органические вещества из неорганических. Для этого они использовали энергию солнечного света. Возникла первая экологическая система, в которой эти организмы были производителями. В результате этого в атмосфере Земли появился кислород, выделяемый этими организмами. С его помощью можно из той же самой пищи получить гораздо больше энергии, а добавочную энергию использовать на усложнение строения тела: разделение тела на части.
Одно из важных достижений жизни — разделение ядра и цитоплазмы. В ядре находится наследственная информация. Специальная мембрана вокруг ядра позволила защитить от случайных повреждений. По мере необходимости цитоплазма получает из ядра команды, направляющие жизнедеятельность и развитие клетки.
Организмы, у которых ядро отделено от цитоплазмы, образовали надцарство ядерных (к ним относятся — растения, грибы, животные).
Таким образом, клетка — основа организации растений и животных — возникла и развилась в ходе биологической эволюции.
Даже не вооружённым глазом, а ещё лучше под лупой можно видеть, что мякоть зрелого арбуза состоит из очень мелких крупинок, или зёрнышек. Это клетки — мельчайшие «кирпичики», из которых состоят тела всех живых организмов, в том числе и растительных.
Жизнь растения осуществляется соединённой деятельностью его клеток, создающих единое целое. При многоклеточности частей растения существует физиологическое разграничение их функций, специализация различных клеток в зависимости от местоположения их в теле растения.
Растительная клетка отличается от животной тем, что имеет плотную оболочку, покрывающую внутреннее содержимое со всех сторон. Клетка не является плоской (как её принято изображать), она скорей всего похожа на очень маленький пузырёк, наполненный слизистым содержимым.
Строение и функции растительной клетки
Рассмотрим клетку как структурно-функциональную единицу организма. Снаружи клетка покрыта плотной клеточной стенкой, в которой имеются более тонкие участки — поры. Под ней находится очень тонкая плёнка — мембрана, покрывающая содержимое клетки — цитоплазму. В цитоплазме есть полости — вакуоли, заполненные клеточным соком. В центре клетки или около клеточной стенки расположено плотное тельце — ядро с ядрышком. От цитоплазмы ядро отделено ядерной оболочкой. По всей цитоплазме распределены мелкие тельца — пластиды.
Строение растительной клетки
Строение и функции органоидов растительной клетки
| Органоид | Рисунок | Описание | Функция | Особенности |
Клеточная стенка или плазматическая мембрана | Бесцветная, прозрачная и очень прочная | Пропускает в клетку и выпускает из клетки вещества. | Клеточная мембрана полупроницаемая |
|
Цитоплазма | Густое тягучее вещество | В ней располагаются все другие части клетки | Находится в постоянном движении |
|
Ядро (важная часть клетки) | Округлое или овальное | Обеспечивает передачу наследственных свойств дочерним клеткам при делении | Центральная часть клетки |
|
Сферической или неправильной формы | Принимает участие в синтезе белка | |||
 | Резервуар, отделённый от цитоплазмы мембраной. Содержит клеточный сок | Накапливаются запасные питательные вещества и продукты жизнедеятельности ненужные клетке. | По мере роста клетки мелкие вакуоли сливаются в одну большую (центральную) вакуоль |
|
Пластиды | Хлоропласты | Используют световую энергию солнца и создают органические из неорганических | Форма дисков, отграниченных от цитоплазмы двойной мембраной |
|
Хромопласты | Образуются в результате накопления каротиноидов | Жёлтые, оранжевые или бурые |
||
 | Лейкопласты | Бесцветные пластиды | ||
Ядерная оболочка | Состоит из двух мембран (наружная и внутренняя) с порами | Отграничивает ядро от цитоплазмы | Даёт возможность осуществляться обмену между ядром и цитоплазмой |
Живая часть клетки — это ограниченная мембраной, упорядоченная, структурированная система биополимеров и внутренних мембранных структур, участвующих в совокупности метаболических и энергетических процессов, осуществляющих поддержание и воспроизведение всей системы в целом.
Важной особенностью является то, что в клетке нет открытых мембран со свободными концами. Клеточные мембраны всегда ограничивают полости или участки, закрывая их со всех сторон.

Современная обобщенная схема растительной клетки
Плазмалемма (наружная клеточная мембрана) — ультрамикроскопическая плёнка толщиной 7,5 нм., состоящая из белков, фосфолипидов и воды. Это очень эластичная плёнка, хорошо смачивающаяся водой и быстро восстанавливающая целостность после повреждения. Имеет универсальное строение, т.е.типичное для всех биологических мембран. У растительных клеток снаружи от клеточной мембраны находится прочная, создающая внешнюю опору и поддерживающая форму клетки клеточная стенка. Она состоит из клетчатки (целлюлозы) — нерастворимого в воде полисахарида.
Плазмодесмы растительной клетки, представляют собой субмикроскопические канальцы, пронизывающие оболочки и выстланные плазматической мембраной, которая таким образом переходит из одной клетки в другую, не прерываясь. С их помощью происходит межклеточная циркуляция растворов, содержащих органические питательные вещества. По ним же идёт передача биопотенциалов и другой информации.
Порами называют отверстия во вторичной оболочке, где клетки разделяют лишь первичная оболочка и срединная пластинка. Участки первичной оболочки и срединную пластинку, разделяющие соседствующие поры смежных клеток, называют поровой мембраной или замыкающей пленкой поры. Замыкающую пленку поры пронизывают плазмодесменные канальцы, но сквозного отверстия в порах обычно не образуется. Поры облегчают транспорт воды и растворенных веществ от клетки к клетке. В стенках соседних клеток, как правило, одна против другой, образуются поры.
Клеточная оболочка имеет хорошо выраженную, относительно толстую оболочку полисахаридной природы. Оболочка растительной клетки продукт деятельности цитоплазмы. В её образовании активное участие принимает аппарат Гольджи и эндоплазматическая сеть.

Строение клеточной мембраны
Основу цитоплазмы составляет ее матрикс, или гиалоплазма, — сложная бесцветная, оптически прозрачная коллоидная система, способная к обратимым переходам из золя в гель. Важнейшая роль гиалоплазмы заключается в объединении всех клеточных структур в единую систему и обеспечении взаимодействия между ними в процессах клеточного метаболизма.
Гиалоплазма (или матрикс цитоплазмы) составляет внутреннюю среду клетки. Состоит из воды и различных биополимеров (белков, нуклеиновых кислот, полисахаридов, липидов), из которых основную часть составляют белки различной химической и функциональной специфичности. В гиалоплазме содержатся также аминокислоты, моносахара, нуклеотиды и другие низкомолекулярные вещества.
Биополимеры образуют с водой коллоидную среду, которая в зависимости от условий может быть плотной (в форме геля) или более жидкой (в форме золя), как во всей цитоплазме, так и в отдельных ее участках. В гиалоплазме локализуются и взаимодействуют между собой и средой гиалоплазмы различные органеллы и включения. При этом расположение их чаще всего специфично для определенных типов клеток. Через билипидную мембрану гиалоплазма взаимодействует с внеклеточной средой. Следовательно, гиалоплазма является динамической средой и играет важную роль в функционировании отдельных органелл и жизнедеятельности клеток в целом.
Цитоплазматические образования – органеллы
Органеллы (органоиды) — структурные компоненты цитоплазмы. Они имеют определённую форму и размеры, являются обязательными цитоплазматическими структурами клетки. При их отсутствии или повреждении клетка обычно теряет способность к дальнейшему существованию. Многие из органоидов способны к делению и самовоспроизведению. Размеры их настолько малы, что их можно видеть только в электронный микроскоп.
Ядро
Ядро — самая заметная и обычно самая крупная органелла клетки. Оно впервые было подробно исследовано Робертом Броуном в 1831 году. Ядро обеспечивает важнейшие метаболические и генетические функции клетки. По форме оно достаточно изменчиво: может быть шаровидным, овальным, лопастным, линзовидным.
Ядро играет значительную роль в жизни клетки. Клетка, из которой удалили ядро, не выделяет более оболочку, перестаёт расти и синтезировать вещества. В ней усиливаются продукты распада и разрушения, вследствие этого она быстро погибает. Образование нового ядра из цитоплазмы не происходит. Новые ядра образуются только делением или дроблением старого.
Внутреннее содержимое ядра составляет кариолимфа (ядерный сок), заполняющая пространство между структурами ядра. В нём находится одно или несколько ядрышек, а также значительное количество молекул ДНК, соединённых со специфическими белками — гистонами.

Строение ядра
Ядрышко
Ядрышко — как и цитоплазма, содержит преимущественно РНК и специфические белки. Важнейшая его функция заключается в том, что в нём происходит формирование рибосом, которые осуществляют синтез белков в клетке.
Аппарат Гольджи
Аппарат Гольджи — органоид, имеющий универсальное распространение во всех разновидностях эукариотических клеток. Представляет собой многоярусную систему плоских мембранных мешочков, которые по периферии утолщаются и образуют пузырчатые отростки. Он чаще всего расположен вблизи ядра.

Аппарат Гольджи
В состав аппарата Гольджи обязательно входит система мелких пузырьков (везикул), которые отшнуровываются от утолщённых цистерн (диски) и располагаются по периферии этой структуры. Эти пузырьки играют роль внутриклеточной транспортной системы специфических секторных гранул, могут служить источником клеточных лизосом.
Функции аппарата Гольджи состоят также в накоплении, сепарации и выделении за пределы клетки с помощью пузырьков продуктов внутриклеточного синтеза, продуктов распада, токсических веществ. Продукты синтетической деятельности клетки, а также различные вещества, поступающие в клетку из окружающей среды по каналам эндоплазматической сети, транспортируются к аппарату Гольджи, накапливаются в этом органоиде, а затем в виде капелек или зёрен поступают в цитоплазму и либо используются самой клеткой, либо выводятся наружу. В растительных клетках Аппарат Гольджи содержит ферменты синтеза полисахаридов и сам полисахаридный материал, который используется для построения клеточной оболочки. Предполагают, что он участвует в образовании вакуолей. Аппарат Гольджи был назван так в честь итальянского учёного Камилло Гольджи, впервые обнаружившего его в 1897 году.
Лизосомы
Лизосомы представляют собой мелкие пузырьки, ограниченные мембраной основная функция которых — осуществление внутриклеточного пищеварения. Использование лизосомного аппарата происходит при прорастании семени растения (гидролиз запасных питательных веществ).

Строение лизосомы
Микротрубочки
Микротрубочки — мембранные, надмолекулярные структуры, состоящие из белковых глобул, расположенных спиральными или прямолинейными рядами. Микротрубочки выполняют преимущественно механическую (двигательную) функцию, обеспечивая подвижность и сокращаемость органоидов клетки. Располагаясь в цитоплазме, они придают клетке определённую форму и обеспечивают стабильность пространственного расположения органоидов. Микротрубочки способствуют перемещению органоидов в места, которые определяются физиологическими потребностями клетки. Значительное количество этих структур расположено в плазмалемме, вблизи клеточной оболочки, где они участвуют в формировании и ориентации целлюлозных микрофибрилл оболочек растительных клеток.

Строение микротрубочки
Вакуоль
Вакуоль — важнейшая составная часть растительных клеток. Она представляет собой своеобразную полость (резервуар) в массе цитоплазмы, заполненную водным раствором минеральных солей, аминокислот, органических кислот, пигментов, углеводов и отделённую от цитоплазмы вакуолярной мембраной — тонопластом.
Цитоплазма заполняет всю внутреннюю полость только у самых молодых растительных клеток. С ростом клетки существенно изменяется пространственное расположение вначале сплошной массы цитоплазмы: у неё появляются заполненные клеточным соком небольшие вакуоли, и вся масса становится ноздреватой. При дальнейшем росте клетки отдельные вакуоли сливаются, оттесняя к периферии прослойки цитоплазмы, в результате чего в сформированной клетке находится обычно одна большая вакуоль, а цитоплазма со всеми органеллами располагаются около оболочки.
Водорастворимые органические и минеральные соединения вакуолей обусловливают соответствующие осмотические свойства живых клеток. Этот раствор определённой концентрации является своеобразным осмотическим насосом для регулируемого проникновения в клетку и выделения из неё воды, ионов и молекул метаболитов.
В комплексе со слоем цитоплазмы и её мембранами, характеризующимися свойствами полупроницаемости, вакуоль образует эффективную осмотическую систему. Осмотически обусловленными являются такие показатели живых растительных клеток, как осмотический потенциал, сосущая сила и тургорное давление.

Строение вакуоли
Пластиды
Пластиды — самые крупные (после ядра) цитоплазматические органоиды, присущие только клеткам растительных организмов. Они не найдены только у грибов. Пластиды играют важную роль в обмене веществ. Они отделены от цитоплазмы двойной мембранной оболочкой, а некоторые их типы имеют хорошо развитую и упорядоченную систему внутренних мембран. Все пластиды едины по происхождению.
Хлоропласты — наиболее распространённые и наиболее функционально важные пластиды фотоавтотрофных организмов, которые осуществляют фотосинтетические процессы, приводящие в конечном итоге к образованию органических веществ и выделению свободного кислорода. Хлоропласты высших растений имеют сложное внутреннее строение.

Строение хлоропласта
Размеры хлоропластов у разных растений неодинаковы, но в среднем диаметр их составляет 4-6 мкм. Хлоропласты способны передвигаться под влиянием движения цитоплазмы. Кроме того, под воздействием освещения наблюдается активное передвижение хлоропластов амебовидного типа к источнику света.
Хлорофилл — основное вещество хлоропластов. Благодаря хлорофиллу зелёные растения способны использовать световую энергию.
Лейкопласты (бесцветные пластиды) представляют собой чётко обозначенные тельца цитоплазмы. Размеры их несколько меньше, чем размеры хлоропластов. Более и однообразна и их форма, приближающая к сферической.

Строение лейкопласта
Встречаются в клетках эпидермиса, клубнях, корневищах. При освещении очень быстро превращаются в хлоропласты с соответствующим изменением внутренней структуры. Лейкопласты содержат ферменты, с помощью которых из излишков глюкозы, образованной в процессе фотосинтеза, в них синтезируется крахмал, основная масса которого откладывается в запасающих тканях или органах (клубнях, корневищах, семенах) в виде крахмальных зёрен. У некоторых растений в лейкопластах откладываются жиры. Резервная функция лейкопластов изредка проявляется в образовании запасных белков в форме кристаллов или аморфных включений.
Хромопласты в большинстве случаев являются производными хлоропластов, изредка — лейкопластов.

Строение хромопласта
Созревание плодов шиповника, перца, помидоров сопровождается превращением хлоро- или лейкопластов клеток мякоти в каратиноидопласты. Последние содержат преимущественно жёлтые пластидные пигменты — каратиноиды, которые при созревании интенсивно синтезируются в них, образуя окрашенные липидные капли, твёрдые глобулы или кристаллы. Хлорофилл при этом разрушается.
Митохондрии
Митохондрии — органеллы, характерные для большинства клеток растений. Имеют изменчивую форму палочек, зёрнышек, нитей. Открыты в 1894 году Р. Альтманом с помощью светового микроскопа, а внутреннее строение было изучено позднее с помощью электронного.

Строение митохондрии
Митохондрии имеют двухмембранное строение. Внешняя мембрана гладкая, внутренняя образует различной формы выросты — трубочки в растительных клетках. Пространство внутри митохондрии заполнено полужидким содержимым (матриксом), куда входят ферменты, белки, липиды, соли кальция и магния, витамины, а также РНК, ДНК и рибосомы. Ферментативный комплекс митохондрий ускоряет работу сложного и взаимосвязанного механизма биохимических реакций, в результате которых образуется АТФ. В этих органеллах осуществляется обеспечение клеток энергией — преобразование энергии химических связей питательных веществ в макроэргиеские связи АТФ в процессе клеточного дыхания. Именно в митохондриях происходит ферментативное расщепление углеводов, жирных кислот, аминокислот с освобождением энергии и последующим превращением её в энергию АТФ. Накопленная энергия расходуется на ростовые процессы, на новые синтезы и т. д. Митохондрии размножаются делением и живут около 10 дней, после чего подвергаются разрушению.
Эндоплазматическая сеть
Эндоплазматическая сеть — сеть каналов, трубочек, пузырьков, цистерн, расположенных внутри цитоплазмы. Открыта в 1945 году английским учёным К. Портером, представляет собой систему мембран, имеющих ультрамикроскопическое строение.
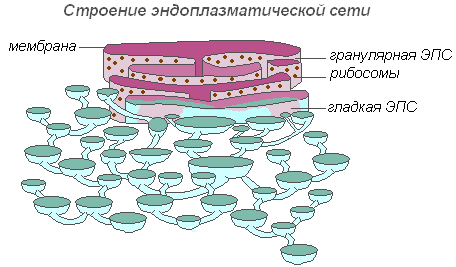
Строение эндоплазматической сети
Вся сеть объединена в единое целое с наружной клеточной мембраной ядерной оболочки. Различают ЭПС гладкую и шероховатую, несущую на себе рибосомы. На мембранах гладкой ЭПС находятся ферментные системы, участвующие в жировом и углеводном обмене. Этот тип мембран преобладает в клетках семян, богатых запасными веществами (белками, углеводами, маслами), рибосомы прикрепляются к мембране гранулярной ЭПС, и во время синтеза белковой молекулы полипептидная цепочка с рибосомами погружается в канал ЭПС. Функции эндоплазматической сети очень разнообразны: транспорт веществ как внутри клетки, так и между соседними клетками; разделение клетки на отдельные секции, в которых одновременно проходят различные физиологические процессы и химические реакции.
Рибосомы
Рибосомы — немембранные клеточные органоиды. Каждая рибосома состоит из двух не одинаковых по размеру частичек и может делиться на два фрагмента, которые продолжают сохранять способность синтезировать белок после объединения в целую рибосому.

Строение рибосомы
Рибосомы синтезируются в ядре, затем покидают его, переходя в цитоплазму, где прикрепляются к наружной поверхности мембран эндоплазматической сети или располагаются свободно. В зависимости от типа синтезируемого белка рибосомы могут функционировать по одиночке или объединяться в комплексы — полирибосомы.
), состоят из большой и малой субъединиц.
Схема синтеза рибосом в клетках эукариот .
1. Синтез мРНК рибосомных белков РНК полимеразой II. 2. Экспорт мРНК из ядра. 3. Узнавание мРНК рибосомой и 4. синтез рибосомных белков. 5. Синтез предшественника рРНК (45S - предшественник) РНК полимеразой I. 6. Синтез 5S pРНК РНК полимеразой III. 7. Сборка большой рибонуклеопротеидной частицы, включающей 45S-предшественник, импортированные из цитоплазмы рибосомные белки, а также специальные ядрышковые белки и РНК, принимающие участие в созревании рибосомных субчастиц. 8. Присоединение 5S рРНК, нарезание предшественника и отделение малой рибосомной субчастицы. 9. Дозревание большой субчастицы, высвобождение ядрышковых белков и РНК. 10. Выход рибосомных субчастиц из ядра. 11. Вовлечение их в трансляцию.
Рибосомы представляют собой нуклеопротеид , в составе которого соотношение РНК/белок составляет 1:1 у высших животных и 60-65:35-40 у бактерий. Рибосомная РНК составляет около 70 % всей РНК клетки. Рибосомы эукариот включают четыре молекулы рРНК, из них 18S , 5,8S и 28S рРНК синтезируются в ядрышке РНК-полимеразой I в виде единого предшественника (45S), который затем подвергается модификациям и нарезанию. 5S рРНК синтезируются РНК-полимеразой III в другой части генома и не нуждаются в дополнительных модификациях. Почти вся рРНК находится в виде магниевой соли, что необходимо для поддержания структуры; при удалении ионов магния рибосома подвергается диссоциации на субъединицы.
РНК большой субъединицы
Высокомолекулярная РНК, составляющая структурную основу большой субъединицы рибосомы, обозначается как 23S рРНК (в случае бактериальных рибосом) или 23S-подобная рРНК (в других случаях). Бактериальная 23S рРНК, также как и 16S рРНК, представляет собой одну ковалентно непрерывную полирибонуклеотидную цепь. В то же время 23S-подобная рРНК цитоплазматических рибосом эукариот состоит из двух прочно ассоциированных полирибонуклеотидных цепей - 28S и 5,8S рРНК (5,8S рРНК является структурным эквивалентом 5′-концевого ~160-нуклеотидного сегмента 23S рРНК, который оказался «отщеплён» в виде ковалентно обособленного фрагмента). 23S-подобная рРНК рибосом пластидов растений также состоит из двух прочно ассоциированных полирибонуклеотидных цепей и содержит 4,5S рРНК - структурный эквивалент 3′-концевого сегмента 23S рРНК. Известны случаи и ещё более глубоко зашедшей фрагментированности РНК, примером чего может служить 23S-подобная рРНК цитоплазматических рибосом некоторых протистов. Так, у Crithidia fasciculata она состоит из 7 отдельных фрагментов, а у Euglena gracilis - из 14.
Кроме вышеуказанной 23S(-подобной) рРНК, большая субъединица обычно содержит также относительно низкомолекулярную РНК - так называемую 5S рРНК. В отличие от вышеупомянутых 5,8S и 4,5S рРНК, 5S рРНК менее прочно ассоциирована с 23S(-подобной) рРНК, транскрибируется с отдельного гена и, таким образом, не может быть рассмотрена как отщеплённый фрагмент высокополимерной рРНК. 5S рРНК входит в состав большой субъединицы цитоплазматических рибосом всех прокариот и эукариот, но, по-видимому, не является непременной составляющей любой функциональной рибосомы, так как 5S рРНК отсутствуют в митохондриальных рибосомах млекопитающих (так называемых «минирибосомах»).
Число нуклеотидных звеньев, как и константы седиментации, для образцов 23S и 23S-подобных рРНК из различных источников могут существенно различаться. Например, 23S рРНК Escherichia coli состоит из 2904 нуклеотидных остатков, цитоплазматическая 26S рРНК Saccharomyces cerevisiae - из 3392, митохондриальная 26S рРНК Saccharomyces cerevisiae - из 3273, цитоплазматическая 28S рРНК Homo sapiens - из 5025. Большие субъединицы митохондриальных рибосом млекопитающих содержат относительно короткие 23S-подобные рРНК - всего 1560-1590 нуклеотидных остатков. Молекула 5,8S рРНК комплекса 28S 5,8S рРНК, характерного для цитоплазматических эукариотических рибосом, имеет длину около 160 нуклеотидных остатков. Длина 5S рРНК довольно консервативна и составляет 115-125 нуклеотидных остатков.
Рибосомные белки
Помимо рРНК, рибосома содержит также около 50 (прокариотические рибосомы) или 80 (цитоплазматические рибосомы эукариот) различных белков . Почти каждый из этих белков представлен лишь одной копией на каждую рибосому. Преобладают умеренно-осно́вные белки. Большинство рибосомных белков эволюционно консервативны, многие белки рибосом из различных источников могут быть соотнесены как гомологи , что учитывается в современной универсальной номенклатуре рибосомных белков. Рибосома на 30-50 % состоит из белка.
Низкомолекулярные компоненты
Кроме биополимеров (РНК и белков) в состав рибосом входят также некоторые низкомолекулярные компоненты. Это молекулы воды, ионы металлов (главным образом Mg 2+ - до 2 % сухой массы рибосомы), ди- и полиамины (такие как путресцин , кадаверин , спермидин, спермин - могут составлять до 2,5 % сухой массы рибосомы).
Механизм трансляции
В 2009 году канадские биохимики Константин Боков и Сергей Штейнберг из Монреальского университета, исследовав третичную структуру рибосомной РНК бактерии Escherichia coli , высказали обоснованное предположение, что рибосомы могли сформироваться в результате постепенной эволюции из очень простой маленькой молекулы РНК - «проторибосомы», способной катализировать реакцию соединения двух аминокислот . Все остальные структурные блоки рибосомы последовательно добавлялись к проторибосоме, не нарушая её структуру и постепенно повышая эффективность её работы .
Примечания
- , с. 109.
- , с. 120-121.
- , с. 110.
- , с. 110-111.
- , с. 133-134.
- , с. 136-137.
- , с. 84-85.
- , с. 84.
- Sievers A. , Beringer M. , Rodnina M. V. , Wolfenden R. The ribosome as an entropy trap. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. - 2004. - 25 May (vol. 101 , no. 21 ). - P. 7897-7901 . - DOI :10.1073/pnas.0402488101 . - PMID 15141076 . [исправить]
В бактериальной рибосомы составляют до 30% ее сухой массы: на
одну бактериальную
приходится примерно 10 4 рибосом. В эукариотич. ( всех
, за исключением бактерий и синезеленых водорослей) относит. содержание
рибосом меньше, и их кол-во очень сильно варьирует в зависимости от белок-синтезирующей
соответствующей или отдельной .
В эукариотич. все
рибосомы (как мембрано-связанные, так и свободные) образуются в ядрышке;
считается, что там они неактивны. Эукариотич. имеет также специальные
рибосомы в (у животных и растений) и (у растений). Рибосомы этих
органелл отличаются от цитоплазматических размерами и нек-рыми функцион. св-вами.
Они образуются непосредственно в этих органеллах.
Различают два осн. типа
рибосом. Всем прокариотич. (бактерии и синезеленые водоросли) свойственны
т. наз. 70S рибосомы, характеризующиеся коэф. () ок. 70 единиц
Сведберга, или 70S (по коэф. различают и рибосомы др. типов, а также
субчастицы и , входящие в состав рибосом). Их мол. м. составляет 2,5 ·
10 6 , линейные размеры 20-25 нм. По хим. составу это рибонуклеопротеиды;
они состоят только из и (соотношение этих компонентов 2:1). Рибосомная
в рибосомах присутствует гл. обр. в виде Mg-соли (по-видимому, частично и в виде
Са-соли); в рибосомах до 2% от сухой массы. Кроме того, в разл. кол-вах (до
2,5%) могут присутствовать также аминов-спермина H 2 N(CH 2) 3 NH(CH 2) 4 NH(CH 2) 3 NH 2 ,
спермидина H 2 N(CH 2) 3 NH(CH 2) 4 NH 2
и др.
По-видимому, определяет
осн. структурные и функцион. св-ва рибосом, в частности обеспечивает целостность рибосомных
субъединиц, обусловливает их форму и ряд структурных особенностей. Специфич.
пространств. структура детерминирует локализацию всех рибосомных ,
играет ведущую роль в организации функцион. центров рибосом.
Рибосомный синтез белка-многоэтапный процесс. Первая стадия (инициация) начинается с присоединения () к малой рибосомной субчастице, не связанной с большой субчастицей. Характерно, что для начала процесса необходима именно диссоциированная рибосома. К образовавшемуся т. наз. инициаторному комплексу присоединяется большая рибосомная субчастица. В стадии инициации участвуют спец. инициирующий (см. ), инициаторная () и специфич. (т. наз. факторы инициации). Пройдя стадию инициации, рибосома переходит к последоват. считыванию по направлению от 5"- к 3"-концу, что сопровождается синтезом полипептидной цепи , кодируемого этой (подробнее о механизме синтеза см. в ст. ). В этом процессе рибосома функционирует как циклически работающая мол. машина. Рабочий цикл рибосомы при состоит из трех тактов: 1) кодонзависимого связывания аминоацил-тРНК (поставляет в рибосому), 2) транспептидации-переноса С-конца растущего на аминоацил-тРНК, т.е. удлинения строящейся белковой цепи на одно звено, 3) транслокации-перемещения () и пептидил-тРНК относительно рибосомы и переход рибосомы в исходное состояние, когда она может воспринять след. аминоацил-тРНК. Когда рибосома достигнет специального , синтез
ПОСМОТРЕТЬ ЕЩЕ:
Рибосома представляет собой элементарную клеточную машину синтеза любых белков клетки. Все они построены в клетке одинаково, имеют одинаковую молекулярную композицию, выполняют одинаковую функцию – синтез белка – поэтому их можно так же считать клеточными органоидами. В отличие от других органоидов цитоплазмы (пластид, митохондрий, клеточного центра, мембранной вакуолярной системы и др.) они представлены в клетке огромным числом: за клеточный цикл их образуется 1 х 107 штук. Поэтому основная масса клеточной РНК представляет собой именно рибосомную РНК. РНК рибосом относительно стабильна, рибосомы могут существовать в клетках культуры ткани в течение нескольких клеточных циклов. В печеночных клетках время полужизни рибосом составляет 50-120 часов.
Рибосомы – это сложные рибонуклеопротеидные частицы, в состав которых входит множество молекул индивидуальных (неповторенных) белков и несколько молекул РНК, Рибосомы прокариот и эукариот по своим размерам и молекулярным характеристикам отличаются, хотя и обладают общими принципами организации и функционирования. К настоящему времени методом рентгеноструктурного анализа высокого разрешения полностью расшифрована структура рибосом.
Полная, работающая рибосома, состоит из двух неравных субъединиц, которые легко обратимо диссоциируют на большую субъединицу и малую. Размер полной прокариотической рибосомы составляет 20 х 17 х 17 нм, эукариотической – 25 х 20 х 20. Полная прокариотическая рибосома имеет коэффициент седиментации 70S и диссоциирует на две субъединицы: 50S и 30S. Полная эукариотическая рибосома, 80S рибосома, диссоциирует на 60S и 40S субъединицы. Форма и детальные очертания рибосом из разнообразных организмов и клеток, включая как прокариотические, так и эукариотические, поразительно похожи, хотя и отличаются рядом деталей. Малая рибосомная субъединица имеет палочковидную форму с несколькими небольшими выступами (см. рис. 81), ее длина составляет около 23 нм, а ширина – 12 нм. Большая субъединица похожа на полусферу с тремя торчащими выступами. При ассоциации в полную 70S рибосому малая субчастица ложится одним концом на один из выступов 50S частицы, а другим в ее желобок. В состав малых субъединиц входит по одной молекуле РНК, а в состав большой – несколько: у прокариот – две, а у эукариот – 3 молекулы. Характеристики молекулярной композиции рибосом даны в таблице 9.
Таблица 9. Молекулярная характеристика рибосом
Таким образом в состав эукариотической рибосомы входят четыре молекулы РНК разной длины: 28S РНК содержит 5000 нуклеотидов, 18SРНК – 2000, 5,8S РНК – 160, 5SРНК – 120.Рибосомные РНК обладают сложной вторичной и третичной структурой, образуя сложные петли и шпильки на комплементарных участках, что приводит к самоупаковке, самоорганизации этих молекул в сложное по форме тело. Так, например, сама по себе молекула 18S РНК в физиологических ионных условиях образует палочковидную частицу, определяющую форму малой субъединицы рибосом.
Под действием низких ионных сил, особенно при удалении ионов магния, плотные рибосомные субъединицы могут разворачиваться в рыхлые рибонуклеопротеидные тяжи, где можно наблюдать кластеры отдельных белков, но правильных структур, типа нуклеосом, нет, т.к. нет групп из сходных белков: в рибосоме все 80 белков разные.
Для того, чтобы образовались рибосомы необходимо наличие четырех типов рибосомных РНК в эквимолярных отношениях и наличие всех рибосомных белков. Сборка рибосом может происходить спонтанно in vitro , если последовательно добавлять к РНК белки в определенной последовательности.
Следовательно для биосинтеза рибосом необходим синтез множества специальных рибосомных белков и 4-х типов рибосомной РНК. Где эта РНК синтезируется, на каком количестве генов, где эти гены локализованы, как они организованы в составе ДНК хромосом – все эти вопросы в последние десятилетия были успешно разрешены при изучении строения и функции ядрышек.
Читайте также:
Рибосомы являются важнейшими органоидами клетки, так как на них протекает процесс трансляции - синтез полипептида на матричной РНК (мРНК). Другими словами, рибосомы служат местом белкового синтеза .
Строение рибосом
Рибосомы относятся к немембранным органоидам. Они очень мелкие (около 20 нм), но многочисленные (тысячи и даже миллионы на клетку), состоят из двух частей – суб ъединиц . В состав субчастиц входят рибосомальные РНК (рРНК) и рибосомные белки, т. е. рибосомы по химическому составу являются рибонуклеопротеи д ами . Однако в них также присутствует небольшое количество низкомолекулярных соединений. Из-за многочисленности рибосом, рРНК составляет более половины от всей РНК клетки.
Одну из субъединиц называют «малой», вторую – «большой».
В собранной из субъединиц рибосоме выделят два (по одним источникам) или три (по другим) участка, которые называют сайтами. Один из участков обозначают A (aminoacyl) и называют аминоацильным, второй - P (peptidyl) - пептидильный. Данные сайты являются основными каталитическими центрами протекающих на рибосомах реакций. Третий участок обозначают E (exit), через него освободившаяся от синтезируемого полипептида транспортная РНК (тРНК), покидает рибосому.
Кроме перечисленных сайтов на рибосомах есть другие участки, используемые для связывания различных ферментов.
Когда субъединицы диссоциированы (разъединены) специфичность сайтов теряется, т. е. они определяются сочетанием соответствующих областей обеих субъединиц.
Отличие рибосом прокариот и эукариот
Соотношение по массе белков и РНК в рибосоме примерно поровну. Однако у прокариот белков меньше (около 40%).
Размеры как самих рибосом, так и субъединиц выражают в скорости их седиментации (осаждения) при центрифугировании. При этом S обозначает константу Сведберга - единицу, характеризующую скорость оседания в центрифуге (чем больше S, тем быстрее частица осаждается, а значит тяжелее). У прокариот рибосомы имеют размер в 70S, а у эукариот - в 80S (т. е. они тяжелее и крупнее). При этом субъединицы прокариотических рибосом имеют значения 30S и 50S, а эукариотических - 40S и 60S. Размеры рибосом в митохондриях и хлоропластах эукариот сходны с прокариотическими (хотя имеют определенную вариабельность по размерам), что может указывать на их происхождение от древних прокариотических организмов.
У прокариот в состав большой субъединицы рибосом входит две молекулы рРНК и более 30 молекул белка, в состав малой - одна молекула рРНК и около 20 белков. У эукариот в субъединицах больше молекул белка, а также в большой субъединице три молекулы рРНК. Составляющие рибосому белки и молекулы рРНК обладают способностью к самосборке и в итоге образуют сложную трехмерную структуру. Структуру рРНК поддерживают ионы магния.
Синтез рРНК
У эукариот в состав рибосом входят 4 вида рРНК. При этом три образуются из одного транскрипта-предшественника - 45S рРНК. Он синтезируется в ядрышке (на петлях хромосом его формирующем) при помощи РНК-полимеразы-1. Гены рРНК имеют много копий (десятки и сотни) и обычно располагаются на концах разных пар хромосом. После синтеза 45S рРНК разрезается на 18S, 5.8S и 28S рРНК, каждая из которых подвергается тем или иным модификациям.
Четвертый вид рРНК синтезируется вне ядрышка с помощью фермента РНК-полимеразы-3. Это 5S РНК, которая после синтеза не нуждается в процессинге.
Третичная структура рРНК в составе рибосом очень сложная и компактная.
Отличия прокариот от эукариот
Она служит каркасом для размещения рибосомных белков, которые выполняют вспомогательные функции для поддержания структуры и функциональности.
Функция рибосом
Функционально рибосомы являются местом связывания молекул, участвующих в синтезе (мРНК, тРНК, различные факторы). Именно в рибосоме молекулы могут занять друг по отношению к другу такое положение, которое позволит быстро протечь химической реакции реакции.
В эукариотических клетках рибосомы могут находиться свободно в цитоплазме или быть прикрепленными с помощью специальных белков к ЭПС (эндоплазматическая сеть, она же ЭР - эндоплазматический ретикулум).
В процессе трансляции рибосома перемещается по мРНК. Часто по одной нитевидной мРНК двигаются несколько (или множество) рибосом, образуя так называемую полисому (полирибосому).
Функциональные центры рибосомы (А-сайт, Р-сайт, ПТФ-сайт, М-сайт, Е-сайт)
Предыдущая12345678910111213141516Следующая
Виды рибосом. Строение рибосомы, малая и большая субъединицы. Состав субъединиц – рибосомальные РНК, рибосомальные белки.
Полные рибосомные частицы и их субъединицы обозначаются в соответствии с коэффициентом седиминтации, выраженном в единицах Сведберга.
Всем прокариотам свойственны 70S рибосомы . Соотношение белка к РНК – 2:1. Состоит из двух субъединиц: 50S и 30S. Каждая содержит рРНК и определенное число небольших белков. У E.coli малая субъединица состоит из 1 рРНК (16S) и 21 рибосомального белка (S1, S2, S3 и тд.). Большая субъединица содержит 2 рРНК (23S, 5S) и 31белок (L1, L2, L3 и тд.). Полная рибосома имеет ассиметричное строение. На малой 4 отдела: гоовка, шейка, тело и основание/платформа. У большой хорошо различим центральный выступ\протурбанец, в кот.находится 5S рРНК, основной массив, в кот. находится белок L7, и бороздка м/д ними, в кот. находится пептидилтрансферазный сайт. М\д большой и малой субъединицей образуется полость, в кот.открывается большинство активных сайтов рибосомы.
Эукариоты имеют 80S рибосомы . Имеют больше рРНК и белков. Их соотношение 1:1. Состоят из малой (40S) и большой (60S) субъединицы. Малая содержит 18SрРНК и 33 рибосомальных белка. Большая – 3 цепи рРНК (5S, 5.8S, 28S) и 45-50 белков.
Рибосомы органелл отличаются от цитомпазматических.
2.2. Рибосомы прокариот и эукариот
Функциональные центры рибосомы (А-сайт, Р-сайт, ПТФ-сайт, М-сайт, Е-сайт).
Рибосома – кооперативная структура, кот.зависит от взаимодействия своих активных сайтов. Сайт А – участвует в связывании очередной аминоацил-тРНК, в нем находится кодон мРНК, кот.диктует рибосоме тип входящей аминоацил-тРНК/следующую аминокислоту растущего полипептида. Сайт Р – участок связывание пептидил-тРНК – растущий петтид, кот. связан своим С-концом с тРНК, кот. принесла последний аминокислотный остаток к рибосоме. Сайт Е – участок выхода тРНК из рибосомы. ДеацелированнаятРНК удерживается сайтом Е короткое время. Уэукариот нет этого сайта, из Р-сайта уходит сразу в цитоплазму. Каталитический сайт пептидилтрансферазы – находится на границе А и Р-сайтов, катализирует образование пептидной связи. ГТФ-азный центр – место посадки ГТФ, содействует запуску гидролиза АТФ
Биосинтез рибосомы,этапы процессинга рРНК. Химические модификации рРНК. Особенности строения и созревания рибосомы эукариот.
Процессинг рРНК: нарезание первичноготранскрипта, метилирование, сплайсинг. Уэукариот все рРНК синтезируются как часть одного транскрипта. Он нарезается с помощью экзо и эндонуклеаз на зрелыерРНК. Предшественник содержит 18, 5.8, 28SрРНК и называется 45S РНК. Процессинг рРНК требует участия мяРНК. У некоторых организмов в составе предшественника 28S РНК находятся вставки/интраны, кот. удаляются в результате процессинга и фрагменты РНК сшиваются в результате сплайсинга.
Упрокариот предшественник рРНК содержит 16, 23, 5SрРНК + несколько предшественников тРНК. 3 и 5’ концы сближены за счет комплиментарно прилегающих пар оснований. Такая структура разрезается РНКазойIII. Оставшиесярибонуклеотиды отрезаются экзонуклеазами/подравнивание.
Предыдущая12345678910111213141516Следующая
Клетки бактерий, сине-зеленых водорослей и актиномицетов содержат рибосомы с коэффициентом седиментации 70S. Этот коэффициент является мерой относительной плавучей плотности частиц при их центрифугировании в градиенте плотности хлористого цезия или сахарозы. Единица плавучей плотности S (сведберг) названа так в честь изобретателя ультрацентрифуги шведского ученого Т. Сведберга. Коэффициент седиментации зависит как от массы, так и от формы частицы. Молекулярная масса прокариотических рибосом составляет 2,5 мД, форма округлая со средним диаметром 25 нм. Общее количество рибосом в бактериальной клетке достигает 30 % ее сухого веса. Относительное количество белка в них в два раза меньше, чем РНК.
Рибосомы прокариотического типа с коэффициентом седиментации 70S содержатся также в хлоропластах высших растений. Однако рибосомы митохондрий, хотя и похожи на бактериальные, обладают более высокой видовой специфичностью. В частности, митохондриальные рибосомы дрожжей несколько крупнее типичных прокариотических рибосом (75S), тогда как митохондриальные рибосомы млекопитающих, наоборот, значительно меньше бактериальных (55S).
Клетки животных, растений, грибов и простейших содержат рибосомы с коэффициентом седиментации 80S. Их молекулярная масса составляет 4 мД, а средний диаметр — 30 нм. Относительное количество белка в них приблизительно равно количеству РНК. Эукариотический тип рибосом не имеет видовых различий.
Морфология рибосом
На маломувеличении электронного микроскопа (до 20 000х) рибосомы выглядят как электронно-плотные округлые частицы диаметром 25-30 нм. На большом увеличении (выше 100 000х) видно, что они разделены бороздкой на две неравные части, представляющие собой малую и большую субъединицы с соотношением масс 1:2.
В физиологических условиях рибосомы обратимо диссоциируют на субъединицы. При этом прокариотические рибосомы диссоциируют по схеме:
70S <=> 30S + 50S,
тогда как эукариотические рибосомы диссоциируют по схеме:
80S <=> 40S + 60S
Дефицит коэффициента седиментации связан с тем, что плавучая плотность рибосом зависит не только от массы субъединиц, но и от их формы.
Малая субъединица прокариотической рибосомы 30S имеет продолговатую форму, ее длина составляет 23 нм, а ширина – 12 нм. Она разделена на доли, которые называются “головка”, “тело” и “боковой выступ”. Наиболее выражена поперечная борозда, которая разделяет головку и тело. Малая субъединица эукариотической рибосомы 40S похожа на малую прокариотическую субъединицу 30S, но имеет две дополнительные детали – выступ головки со стороны, противоположной боковому выступу тела, а также раздвоенность дистального конца тела.
Большая субъединица прокариотической рибосомы 50S диаметром 25 нм внешне идентична большой субъединице эукариотической рибосомы 60S. В большой субъединице имеются три выступа: средний выступ или “головка”, боковая доля или “ручка”, палочковидный отросток или “носик”. В целом форма большой субъединицы напоминает чайник для заварки.
Объединение субъединиц в полную рибосому происходит строго закономерным образом. При этом головки и боковые выступы малой и большой субъединиц ориентируются в одну сторону и накладываются друг на друга. Уплощенные поверхности субъединиц также взаимно дополняют друг друга в пространстве.
Химический состав рибосом
Рибосома состоит из РНК и белков, причем основные структурно-функциональные свойства этого органоида определяются рибосомальной РНК.
Прокариотические рибосомы содержат три, а эукариотические — четыре молекулы рибосомальной РНК.
Рибосомальные РНК
РНК малой субъединицы с коэффициентами седиментации 16S и 18S имеет от 1500 до 1800 нуклеотидных остатков. Она обладает значительной внутренней комплементарностью, за счет чего формируется около трех десятков коротких двуспиральных участков – “шпилек”, которые детерминируют форму малой субчастицы.
Длинная молекула РНК большой субъединицы с коэффициентом седиментации 18S или 26S содержит от 3000 до 4800 нуклеотидных остатков. За счет внутренней комплементарности в ней формируется более 100 двойных спиралей, которые определяют форму субъединицы.
Кроме длинной РНК, большая субъединица прокариотических и эукариотических рибосом содержит также короткую 5S РНК, состоящую из 120 нуклеотидных остатков, которая за счет внутренней комплементарности формирует Т-образную структуру с 5 спиральными участками.
Большая субъединица эукариотических рибосом содержит дополнительно 5,8S РНК.
Рибосомы прокариот и эукариот
Она состоит из 160 нуклеотидных остатков и комплементарно связана с 26S РНК. Следует отметить, что 5,8S РНК большой субъединицы эукариотических рибосом гомологична 5’-концу бактериальной 23S РНК.
Таким образом, основная функция рибосомальных РНК состоит в формировании молекулярного скелета малой и большой субъединиц рибосомы.
Рибосомы содержат 50-70 различных белков, причем большинство из них представлено лишь одной молекулой. Молекулярная масса рибосомальных белков находится в пределах 10-30 кД, хотя отдельные полипептиды достигают массы 70 кД. Среди рибосомальных белков преобладают основные полипептиды, но встречаются также нейтральные и кислые белки. Малая субъединица прокариотической рибосомы содержит 20 белков, а большая – 30 белков. У эукариотических рибосом белков значительно больше: малая субъединица содержит 30 белков, а большая — 40.
Рибосомальные белки осуществляют разнообразные функции, связанные с ролью рибосомы как организатора биосинтеза белка:
- формируют участки малой и большой субъединиц;
- образуют центры связывания молекул;
- катализируют химические реакции;
- участвуют в регуляции биосинтеза белка;
Многие рибосомальные белки выполняют одновременно несколько функций.
Белоксинтезирующая система
Наследственная информация закодирована в первичной структуре ДНК, которая в эукариотических клетках сосредоточена в клеточном ядре. Участки ДНК, кодирующие первичную структуру полипептида – структурные гены, являются матрицами для синтеза информационной РНК (иРНК). Процесс образования функциональных копий генов в виде иРНК называется транскрипцией .
Отредактированные в ходе сплайсинга иРНК поступают затем в цитоплазму, где связываются с рибосомами. Используя информацию, закодированную в иРНК, рибосомы синтезируют полипептид в ходе процесса, называемого трансляцией . Синтез полипептида из аминокислот осуществляется в соответствии с генетическим кодом , который представляет собой правила соответствия аминокислот триплетам нуклеотидов в иРНК (кодонам ).
Кроме иРНК и рибосом для осуществления трансляции необходим еще ряд других молекул. Рибосомы совместно с молекулами, принимающими участие в трансляции, образуют белоксинтезирующую систему , которая может функционировать вне клетки. Составы минимальной и полной бесклеточной систем трансляции на прокариотических рибосомах представлены в следующей таблице.
Рибосомы эукариот и прокариот, сходства и различия
Рибосома (от «РНК» и soma – тело) – клеточный немембранный органоид, осуществляющий трансляцию (считывание кода мРНК и синтез полипептидов). Молекула белка рождается в цитоплазме клетки на свободных рибосомах или на цистернах транспортно-накопительной системы. Специальные белки шапероны, укладывают растущую цепочку в ажурную конструкцию. Затем, если нужно, белок достраивают. Различают 2 основных типа рибосом – прокариотные и эукариотные. В митохондриях и хлоропластах также имеются рибосомы, которые близки к рибосомам прокариот. Рибосомы эукариот расположены на мембранах эндоплазматической сети (гранулярная ЭС) и в цитоплазме. Прикрепленные к мембранам рибосомы синтезируют белок «на экспорт», а свободные рибосомы – для нужд самой клетки.
Рибосомы прокариот
У прокариот генетический материал не изолирован от аппарата трансляции, и прокариотные рибосомы занимают почти весь цитоплазматический компартмент. Относительное (по сравнению с другими органеллами) количество рибосом у прокариотов выше, чем у эукариотов, и это обеспечивает более высокую активность их метаболизма, а также более высокую скорость их роста и размножения. Рибосомы - это множественные генеральные микрокомпартменты, которые находятся в цитоплазматическом компартменте и выполняют роль универсальной белоксинтезирующей органеллы. За исключением редких случаев, когда полипептиды синтезируются нерибосомным путем, аминокислоты связываются в линейную цепь только благодаря ферментативной активности рибосом.
Биосинтетический процесс трансляции уникален, поскольку информация о порядке расположения нуклеотидных триплетов (код мРНК) переводится в информацию о порядке расположения аминокислот (код полипептида). Посредником между этими кодами, или «адаптером» (англ. adapter - звукосниматель аудиосистемы) является тРНК. Она доставляет аминокислоту в пептидилтрансферазный центр и одновременно с этим распознает ее кодон в молекуле мРНК.
Типы рибосом. Рибосома - это мультимолекулярный комплекс, состоящий из рРНК и рибосомных белков в массовом соотношении 2:1. В рабочем состоянии рибосома, или «моносома» представляет собой частицу диаметром 25 нм, которая состоит из двух субъединиц - большой L-субъединицы (от англ. large) и малой S-субъединицы (от англ. small). Они имеют разный состав, разную морфологию и выполняют разные функции.
По количественным признакам все рибосомы подразделяются на два типа - прокариотный и эукариотный. Прокариотная рибосома имеет коэффициент седиментации 70S (субъединицы 50S и 30S), а эукариотная рибосома -80S (субъединицы 60S и 40S). Прокариотная рибосома содержит три молекулы рРНК - 23S (~3000 нуклеотидов), 16S (~1500 нуклеотидов) и 5S (~120 нуклеотидов), а также 53-65 однокопий-ных белков. Эукариотная рибосома устроена сложнее прокариотной. Она содержит не три, а четыре молекулы рРНК -28S (4000-6000 нуклеотидов), 18S (1750-1850 нуклеотидов), 5S (~120 нуклеотидов) и 5,8S (~150 нуклеотидов), а также более богатый набор однокопийных белков (70-84).
Механизм трансляции.
Хотя современные представления об архитектуре рибосом и процессе трансляции сложились на основе данных, полученных на бактериях, доказано, что работа рибосомы универсальна у всех трех глобальных доменов. Результаты рентген-структурного анализа с уровнем разрешения 5,5А и криоэлектронной микроскопии дали для рибосомы Е. coli картину геометрического тела сложнейшей конфигурации, состоящего из взаимно переплетенных молекул рРНК и белка. Внутри него, а также на его поверхности имеются каналы, борозды, углубления, площадки, выступы и мостики.
Субстратами для биосинтеза полипептидной цепи служат аминоацил-тРНК, причем для каждой аминокислоты существует своя тРНК и своя аминоацил-тРНК-синтетаза. Специфические тРНК (~75 нуклеотидов) различаются по первичной структуре, однако все они имеют стандартную Г-образную третичную структуру. На дистальном конце длинного «локтя» находится антикодон, комплементарный триплету мРНК, который кодирует специфическую аминокислоту. На дистальном конце короткого «локтя» всех тРНК находится 3′-концевая последовательность ССА. К аденозину (по его 2′- или З’-гидроксильному радикалу) присоединяется а-карбоксильная группа специфической аминокислоты. В ходе трансляции антикодон длинного «локтя» распознает на S-субъединице кодон мРНК, а короткий «локоть» с аминокислотой взаимодействует на L-субъединице с пептидилтрансферазным центром, который катализирует образование пептидной связи.
Долгое время считали, что трансляцию обеспечивают рибосомные белки, а рРНК служит лишь каркасом для их сборки. Однако в настоящее время доказано, что роль главного катализатора трансляции играет рРНК, а белки выполняют структурную функцию.
Между субъединицами рибосомы существует разделение труда. Малая субъединица содержит декодирующий центр, который обеспечивает взаимодействие между мРНК и тРНК. Большая субъединица содержит пептидилтрансферазный центр. В организации декодирующего центра участвуют 16S рРНК и рибосомные белки, в то время как пептидилтрансферазный центр образован только 23S рРНК. Последовательность Шайна-Дальгарно (J. Shine, L. Dalgarno), предшествующая стартовому кодону мРНК, спаривается с комплементарной последовательностью на З’-конце 16S рРНК. Антикодоновый конец тРНК также взаимодействует с 16S рРНК, тогда как акцепторный конец тРНК взаимодействует с 23S рРНК.
Рибосома шаг за шагом образует пептидные связи в направлении от N-конца к С-концу. Для инициации полипептидной цепи у бактерий используется особая аминокислота - формилметионин, которая доставляется в рибосому с помощью специфической тРНК. Происходит спонтанная пептидилтрансферазная реакция: нуклеофильная а-аминогруппа аминоацил-тРНК атакует электрофильную карбонильную группу (*) в сложной эфирной связи между пептидом (или формил-метиониновой затравкой) и другой тРНК. Молекулы 16S рРНК и 23S рРНК образуют три сайта -Р, А и Е, каждый из которых представлен субсайтами в обеих субъединицах рибосомы. P-сайт (от англ. peptide) связывает пептидил-тРНК, A-сайт (от англ. amino acid) связывает аминоацил-тРНК, а Е-сайт (от англ. exit) связывает деацилированную тРНК.
Рабочий цикл рибосомы состоит из четырех этапов, или состояний.
1. В исходном состоянии Р/Р-А/А пептидил-тРНК находится в P-сайте, аминоацил-тРНК в A-сайте, Е-сайт свободен. Специфическая аминоацил-тРНК связывается с A-сайтом при помощи фактора элонгации EF-Tu. Для этого используется энергия гидролиза ГТФ. Тройственный комплекс аминоацил-тРНК/(ЕГ-Ти) × ГТФ прочно связывается с рибосомой только если антикодон комплементарен кодону в декодирующем субсайте А.
2. В «претранслокационном» состоянии Р/Р-А/А происходит пептидилтрансфераз-ная реакция. При этом аминокислота, находящаяся в A-сайте, образует связь с пептидом (или формилметионином, если цепь инициируется), который находится в P-сайте. В обоих случаях дипептид или полипептидная цепь, удлиннившаяся на один аминокислотный остаток, переносятся в А-сайт. Чтобы следующая молекула аминоацил-тРНК могла попасть в A-сайт, пептидил-тРНК должна освободить его и перейти в P-сайт. Этот процесс называется «транслокацией». При транслокации взаимодействующие друг с другом тРНК и мРНК перемещаются внутри рибосомы на расстояние до 50А.
3. В «гибридном транслокационном» состоянии Е/Р-Р/А связанный с пептидом конец тРНК перемещается на большой субъединице из А-субсайта в Р-субсайт, а акцепторный ССА-конец деацилированной тРНК перемещается из Р-субсайта в Е-субсайт. Этот этап транслокации напоминает «эффект домино» и зависит от рРНК. Его механизм еще неизвестен.
4. В «однородном транслокационном» состоянии Е/Е-Р/Р антикодоновый конец тРНК, связанной с пептидом, перемещается на малой субъединице из А-субсайта в ее же Р-субсайт, а антикодоновый конец деацилированной тРНК перемещается из Р-субсайта в Е-субсайт. В результате этого мРНК сдвигается в малой субъединице на один кодон. Теперь A-сайт может принять следующую молекулу аминоацил-тРНК, а деацилированная молекула тРНК может покинуть рибосому. Хотя этот этап транслокации зависит от рРНК, он ускоряется фактором элонгации EF-G, который использует энергию гидролиза ГТФ.
Действие многих антибиотиков (канамицина, неомицина, олеандомицина, стрептомицина, тетрациклина, хлорамфеникола и др.) основано на их связывании с факторами элонгации и сайтами, которые образует рРНК.
Структура рибосом эукариот
Рибосомы состоят из двух различных субчастиц, каждая из которых построена из рибосомной РНК и многих белков. Рибосомы и их субчастицы обычно классифицируют не по массам, а в соответствии с коэффициентами седиментации. Так. коэффициент седиментации полной эукариотической рибосомы составляет около 80 единиц Сведберга (80S), а коэффициент седиментации ее субчастиц составляет 40S и 60S.
Меньшая 40S-субчастица состоит из одной молекулы 18S-рРНК и 30-40 белковых молекул. Большая 60S-субчастица содержит три типа рРНК с коэффициентами седиментации 5S, 5,8S и 28S и 40-50 белков (например, рибосомы гепатоцитов крысы включают 49 белков). В присутствии мРНК (mRNA) субчастицы объединяются с образованием полной рибосомы, масса которой примерно в 650 раз больше массы молекулы гемоглобина. Рибосомы имеют диаметр 20-200 нм и их можно видеть в электронный микроскоп. Структурная организация рибосом полностью не выяснена. Однако известно, что молекулы мРНК проходит через щель около характерной структуры в виде «рога» на малой субчастице, причем эта щель ориентирована как раз в промежуток между двумя субчастицами. тРНК также связываются вблизи этого участка. Для сравнения на схеме в том же масштабе показана молекула тРНК.
В клетках эукариот рибосомы формируются в ядрышке, где на ДНК синтезируется р-РНК, к которой затем присоединяются белки. Субчастицы рибосомы выходят из ядра в цитоплазму, и здесь завершается формирование полноценных рибосом. В цитоплазме рибосомы свободно находятся в цитоплазматическом матриксе (гиалоплазме) или прикрепляются к внешним мембранам ядра и эндоплазматической сети. Свободные рибосомы синтезируют белки для внутренних нужд клетки. Рибосомы на мембранах образуют комплексы – полирибосомы, которые синтезируют белки, поступающие через эндоплазматическую сеть в аппарат Гольджи и затем секретируемые клеткой. Количество рибосом в клетке зависит от интенсивности биосинтеза белка – их больше в клетках активно растущих тканей (меристем растений, зародышей и т. п.). В хлоропластах и митохондриях есть свои собственные мелкие рибосомы, они обеспечивают этим органоидам автономный (независимый от ядра) биосинтез белков.
Каждая рибосома состоит из двух субчастиц — большой и малой. Рибосомы состоят из примерно равных (по массе) количеств РНК и белка (т.е. представляют собой рибонуклеопротеиновые частицы). Входящая в их состав РНК, называемая рибосомной РНК (рРНК), синтезируется в ядрышке.
Вместе те и другие образуют сложную трехмерную структуру, обладающую способностью к самосборке. Во время синтеза белка на рибосомах аминокислоты, из которых строится полипептидная цепь, последовательно одна за другой присоединяются к растущей цепи. Рибосома служит местом связывания для молекул, участвующих в синтезе, т. е. таким местом, где эти молекулы могут занять по отношению друг к другу совершенно определенное положение.
В синтезе участвуют: матричная РНК (мРНК), несущая генетические инструкции от ядра клетки, транспортная РНК (тРНК), доставляющая к рибосоме требуемые аминокислоты, растущая полипептидная цепь, а также ряд факторов, ответственные за инициацию, элонгацию и терминацию цепи. В эукариотических клетках отчетливо видны две популяции рибосом — свободные рибосомы и рибосомы, присоединенные к эндоплазматическому ретикулуму. Строение тех и других идентично, но часть рибосом связана с эндоплазматическим ретикулоумом через белки, которые они синтезируют. Такие белки обычно секретируются. Примером белка, синтезируемого свободными рибосомами, может служить гемоглобин, образующийся в молодых эритроцитах. В процессе синтеза белка рибосома перемещается вдоль нитевидной молекулы мРНК. Процесс идет более эффективно, когда вдоль мРНК перемещается не одна рибосома, а одновременно много рибосом, напоминающих в этом случае бусины на нитке. Такие цепи рибосом называются полирибосомами или полисомами. На эндоплазматическом ретикулуме полисомы обнаруживаются в виде характерных завитков.
Рибосомный синтез белка-многоэтапный процесс. Первая стадия (инициация) начинается с присоединения матричной РНК (мРНК) к малой рибосомной субчастице, не связанной с большой субчастицей. Характерно, что для начала процесса необходима именно диссоциированная рибосома. К образовавшемуся т. наз. инициаторному комплексу присоединяется большая рибосомная субчастица. В стадии инициации участвуют спец. инициирующий кодон (см. Генетический код), инициаторная транспортная РНК (тРНК) и специфич. белки (т. наз. факторы инициации). Пройдя стадию инициации, рибосома переходит к последоват. считыванию кодонов мРНК по направлению от 5′- к 3′-концу, что сопровождается синтезом полипептидной цепи белка, кодируемого этой мРНК В этом процессе рибосома функционирует как циклически работающая мол. машина.
Рабочий цикл рибосомы при элонгации состоит из трех тактов: 1) кодонзависимого связывания аминоацил-тРНК (поставляет аминокислоты в рибосому), 2) транспептидации-переноса С-конца растущего пептида на аминоацил-тРНК, т.е. удлинения строящейся белковой цепи на одно звено, 3) транслокации-перемещения матрицы (мРНК) и пептидил-тРНК относительно рибосомы и переход рибосомы в исходное состояние, когда она может воспринять след. аминоацил-тРНК. Когда рибосома достигнет специального терминирующего кодона мРНК, синтез полипептида прекращается. При участии специфич. белков (т. наз. факторов терминации) синтезир. полипептид освобождается из рибосомы. После терминации рибосома может повторить весь цикл с др. цепью мРНК или др. кодирующей последовательностью той же цепи.
В клетках с интенсивной секрецией белка и развитым эндоплазматич. ретикулумом значит. часть цитоплазматической рибосомы прикреплена к его мембране на поверхности, обращенной к цитоплазме. Эти рибосомы синтезируют полипептиды, которые непосредственно транспортируются через мембрану для дальнейшей секреции. Синтез полипептидов для внутриклеточных нужд происходит в основном на свободных (не связанных с мембраной) рибосомах цитоплазмы. При этом транслирующие рибосомы не равномерно диспергированы в цитоплазме, а собраны в группы. Такие агрегаты рибосом представляют собой структуры, где мРНК ассоциирована со многими рибосомами, находящимися в процессе трансляции; эти структуры получили назв. полирибосом или полисом.
При интенсивном синтезе белка расстояние между рибосомами вдоль цепи мРНК в полирибосоме может быть предельно коротким, т.е. рибосомы находятся почти вплотную друг к другу. Рибосомы, входящие в полирибосомы, работают независимо и каждая из них синтезирует полную полипептидную цепь.
Отличия в строении рибосом прокариотов и эукариотов
Прокариотическая клетка содержит несколько тысяч рибосом, в эукариотической клетке их в десятки раз больше. Рибосомы про- и эукариот отличаются по размерам (у прокариот они мельче, чем у эукариот), но принцип их строения одинаков. Состоят рибосомы из двух частей: большой и малой субъединиц. В их состав кроме белков входят РНК. Эти РНК получили название рибосомных, рРНК.
Величину рибосом и составляющих их частей принято указывать в специальных единицах — S (Сведберг). S — это коэффициент седиментации, который характеризует скорость перемещения молекул или частиц в центробежном поле при центрифугировании. Скорость перемещения зависит от массы частиц, их размеров и формы. Величина рибосом прокариот и эукариот — 70S и 80S соответственно.
В рибосомы прокариот входит три разных вида молекул рРНК (16S рРНК — в малую; 23S рРНК и 5S рРНК — в большую субъединицы) и 55 различных белков (21 — в малую и 34 — в большую субъединицы). В состав эукариотических рибосом входят четыре вида молекул рРНК (18S рРНК — в малую; 28S рРНК, 5.8S рРНК и 5S рРНК — в большую субъединицы) и около 80 белков. В митохондриях и хлоропластах также обнаружены рибосомы. Они характеризуются теми же свойствами и параметрами, что и рибосомы прокариот.
Молекулы рРНК взаимодействуют друг с другом и с белками, образуя компактные структуры — субъединицы рибосом. У эукариот соединение рРНК с рибосомными белками происходит в ядрышке. В центре ядрышка расположен участок хромосомы, в котором находятся гены рибосомных РНК. Синтезированные рРНК соединяются с рибосомными белками, которые поступили через ядерные поры из цитоплазмы, где они были синтезированы на уже существовавших рибосомах. Они соединяются с молекулами рРНК, образуя субъединицы рибосом. Готовые субъединицы через поры выходят в цитоплазму, где будут участвовать в синтезе белка.
Таким образом, ядрышко — это не только место синтеза рибосомных РНК, но и место сборки субъединиц рибосом. Рибосомы нужны в огромных количествах, поскольку в клетке постоянно идут процессы синтеза белка. Поэтому на хромосомах в тех местах, где расположены гены рРНК, находится громадное скопление молекул: синтезируемые рРНК, пришедшие из цитоплазмы рибосомные белки, собираемые и готовые суъединицы рибосом. Понятно, почему ядрышко является самой плотной частью ядра и клетки. Размеры ядрышка зависят от функционального состояния клеток. Если в клетке активно идут процессы биосинтеза белков, ядрышко может занимать до 25% от объема ядра.
Ядрышко образуется на тех хромосомах, где есть гены рРНК. Эти участки хромосом называются ядрышковыми организаторами. Например, у человека десять хромосом способны образовывать ядрышки. Каждый ядрышковый организатор представляет собой огромную хроматиновую петлю, так как содержит несколько десятков и даже сотен одинаковых последовательностей — генов рРНК. Эти последовательности расположены друг за другом и синтез рРНК идет одновременно со всех копий. Таким образом увеличивается интенсивность синтеза рРНК, на долю которой приходится более 90 % всей РНК клетки. Ядрышки, образованные разными хромосомами, очень часто сливаются друг с другом. В ядрах клеток человека обычно наблюдают одно, два или три ядрышка.
При начале трансляции малая субъединица рибосомы связывается с определенным участком иРНК, к ним присоединяется тРНК с аминокислотой, а затем с этим комплексом связывается большая субъединица. После этого рибосома готова к выполнению своей функции — синтезу белка. Белки рибосом способны выполнять свои функции только в составе рибосомы -только в комплексе с рРНК и другими рибосомными белками они приобретают небходимую конформацию.
Эукариотная транскрипция разделена с трансляцией в пространстве и времени. Транскрипция вместе с процессингом РНК происходят в нуклеоплазме, а трансляция, в зависимости от типа клеток, осуществляется преимущественно в цитозоле или на шероховатом эндогшазматическом ретикулуме (англ. rough endoplasmic reticulun, RER). Интегральные белки встраиваются в мембрану RER котрансляционно, а секретируемые белки выделяются в полость цистерны RER через тороидальный переходник между выходным порталом рибосомы и мембранным транслоконом (его образует белок Sec61).
У прокариотов не существует пространственно-временной изоляции процессов транскрипции и трансляции. Цитоплазматические рибосомы присоединяются к 5′-концу мРНК еще до завершения образования короткоживущего транскрипта. Котрансляционная инсерция интегральных белков известна только на примере «шероховатых тилакоидов» цианобактерий. Гидрофобные белки при помощи SRP-частиц презентируются транс локону - компоненту генеральной системы секреции Sec.
Транспортная РНК, напоминает в развернутой форме клеверный лист. Аминокислота прикреплена к “черешку клеверного листа”, а на вершине листа находится триплет, взаимодействующий с кодоном в иРНК — антикодон. Роль "заглавной буквы" при трансляции аминокислотной последовательности у прокариот выполняет измененная форма аминокислоты метионина — формилметионин. Ей соответствует кодон АУГ. После завершения синтеза полипептидной цепи формилметионин отщепляется и в готовом белке отсутствует. В том случае, когда триплет АУГ стоит внутри гена, он кодирует неизмененную аминокислоту метионин.
Если кодон и антикодон комплементарны друг другу, то рибосома передвигается относительно иРНК, и следующий кодон становится доступным для взаимодействия со следующей тРНК. Происходит отсоединение первой аминокислоты от первой тРНК и присоединение ее к аминокислоте, которую принесла вторая тРНК. Во время передвижения рибосомы относительно иРНК первая тРНК, свободная от аминокислоты, покидает рибосому. Вторая тРНК остается, соединенная с пептидом из двух аминокислотных остатков, и в рибосому входит третий кодон иРНК для взаимодействия с очередной тРНК и т.д.
Когда в рибосоме оказывается один из трех триплетов (УАА, УАГ, УГА), ни одна тРНК не может занять место напротив него, так как не существует тРНК с антикодонами, комплементарными этим последовательностям. Полипептидной цепи не к чему присоединиться и она покидает рибосому. Синтез белка завершен. Таким образом, рибосома соединяет в одном месте участников трансляции: иРНК и аминокислоты в комплексе с тРНК, при этом молекулы РНК так ориентированы относительно друг друга, что становится возможным кодон-антикодоновое взаимодействие. Образование пептидной связи контролируется правильностью кодон-антикодонового взаимодействия. Рибосома осуществляет образование пептидной связи и перемещение относительно иРНК.
Молекула информационной РНК взаимодействует не с одной рибосомой, а с несколькими. Каждая рибосома проходит весь путь от "заглавного" кодона до терминирующего, синтезируя одну молекулу белка. Чем больше рибосом пройдет по иРНК, тем больше молекул белка будет синтезировано. Молекула информационной РНК с несколькими рибосомами похожа на нитку бус и называется полирибосомой, или полисомой.